Hepatitis C Virus
Life Cycle
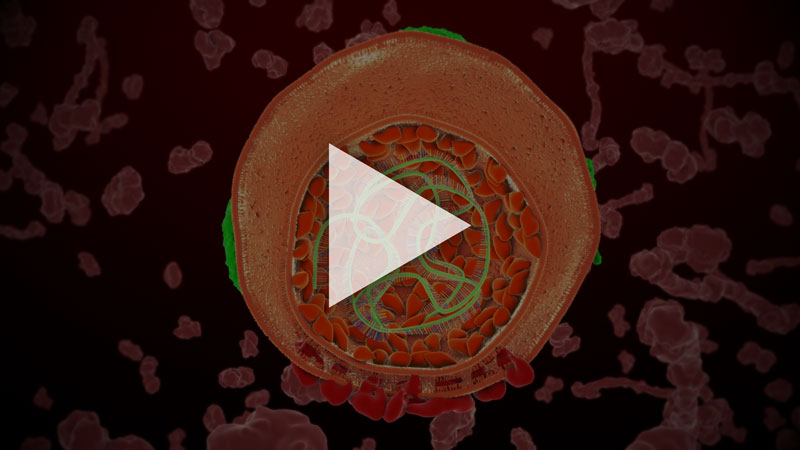
L'hépatite C est une maladie infectieuse transmissible par le sang et due au virus de l'hépatite C (VHC) qui se multiplie dans les cellules du foie. Contrairement aux hépatites virales A et B, il n'existe pas de vaccin contre ce virus. Environ 80 % des gens contaminés par le virus ne parviennent pas à s'en débarrasser spontanément. Dans ce cas, l'hépatite C devient chronique. Dans 20 % des cas, elle évolue vers une cirrhose, 10 a 20 ans après la contamination. Dans 3 à 10 % des cas, cette cirrhose entraîne un cancer du foie (carcinome hépatocellulaire). Le traitement actuel est une bithérapie interféron-ribavirine coûteuse dont l'efficacité est très relative et entraine de nombreux effets secondaire. L'hépatite C est actuellement la cause majeure des transplantation hépatique.
Identifié en 1989 (1), le VHC est un petit virus enveloppé d'environ 60 nanomètres de diamètre, possédant une enveloppe contenant la capside protéique que l'on suppose icosaèdrique. Son génome est un ARN monocaténaire linéaire de polarité positive qui, associé à la protéine de capside, forme la nucléocapside. Il existe sept génotypes du virus de l'hépatite C et de nombreux sous types indiqués par un nombre et une lettre. Le genotype 1b est le plus difficile à soigner.
Le VHC est classé dans la famille des Flaviviridae qui se compose de trois genres :
La structure du VHC reste encore à élucider. Les particules ont un diamètre de 40 à 70 nm (2). La protéine core et les glycoprotéines d'enveloppe E1 et E2 composent le virion. On suppose que E1 et E2 sont ancrées dans une bicouche lipidique entourant la nucléocapside, composée de multiple copies de la protéine core, et de l'ARN génomique.
Le VHC circule sous diverses formes chez un hôte infecté, soit sous forme libre, soit associé à des lipoprotéines de faible densité (LDL) et de très faible densité (VLDL), ceux-là représentant la fraction infectieuse majeure du virus (3).
Le VHC n'infecte que les humains, et le chimpanzé en laboratoire. Les hépatocytes sont la cible principale du virus mais ce dernier est également capable d'infecter entre autres les cellules B et les cellules denditriques.
Les récepteurs cellulaires au virus, supposés ou identifiés, sont:
Bien que son rôle exact reste à déterminer, le récepteur aux LDL (LDLR) jouerait un rôle important du fait de l'association du virus aux LDL et VLDL. La dynamique d'interaction reste mal connue. Avec les glycosaminoglycanes (GAG), le LDLR et d'autres protéines de surface impliquées dans la liaison avec les lipoprotéines et leur métabolisme pourrait servir de collecteur du VHC, facilitant ensuite sa liaison aux autres récepteurs de surface.
Les recepteurs SR-BI et la CD81 se liraient toutes deux à la glycoprotéine E2. Ils sont nécessaires mais pas suffisants à l'entrée du virus. La Claudine-1, récemment identifiée comme co-récepteur au VHC (7), est essentielle à l'entrée du virus.
Tout comme l'occludine, elle agit à une étape tardive de l'entrée, après reconnaissance de CD81.
L'entrée du VHC dans la cellule se ferait par endocytose clathrine-dépendante (9).
Dans l'endosome, l'acidification progressive (10, 11) induirait des changements conformationnels des glycoprotéines d'enveloppe, aboutissant à la fusion entre membranes virale et endosomale (12, 13) et à la libération de la nucléocapside dans le cytosol.
1. Choo, Q.-L. et al. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 244, 359–362 (1989).
2. Wakita, T. et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nature Med. 11, 791–796 (2005).
3. André, P. et al. Hepatitis C virus particles and lipoprotein metabolism. Semin. Liver Dis. 25, 93–104 (2005).
4. Pileri, P. et al. Binding of hepatitis C virus to CD81. Science 282, 938–941 (1998).
5. Agnello, V., Abel, G., Elfahal, M., Knight, G. B. & Zhang,Q. X. Hepatitis C virus and other flaviviridae viruses enter cells via low density lipoprotein receptor. Proc. Natl. Acad. Sci. USA 96, 12766–12771 (1999).
6. Scarselli, E. et al. The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus. EMBO J. 21, 5017–5025 (2002).
7. Evans, M. J. et al. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature 446, 801–805 (2007).
8. Ploss, A. et al. Human occludin is a hepatitis C virus entry factor required for infection of mouse cells. Nature 457, 882-886 (2009)
9. Blanchard, E. et al. Hepatitis C virus entry depends on clathrin-mediated endocytosis. J. Virol. 80, 6964–6972 (2006)
10. Koutsoudakis, G. et al. Characterization of the early steps of hepatitis C virus infection by using luciferase reporter viruses. J. Virol. 80, 5308–5320 (2006).
11. Tscherne, D. M. et al. Time- and temperaturedependent activation of hepatitis C virus for low-pH triggered entry. J. Virol. 80, 1734–1741 (2006)
12. Haid S et al. Low pH-dependent hepatitis C virus membrane fusion depends on E2 integrity, target lipid composition, and density of virus particles. J Biol Chem. 284, 17657-17667 (2009)
13. Lavillette D et al. Hepatitis C virus glycoproteins mediate low pH-dependent membrane fusion with liposomes. J Biol Chem. 281, 3909-3917 (2006 )